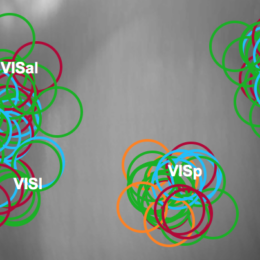
Allen Institute’s Brain Observatory data set
Posted in Uncategorized
The Allen Institute has released the first set of data from their Brain Observatory project. Many of you already know about this, but I wanted to post about it to encourage people to take a…








{kind=link}
{kind=link}